
New Discoveries About How Your Brain Cleans House While You Sleep
- Fred Shaffer
- Apr 7
- 15 min read


Overview
We have long told patients that sleep is restorative, but until recently, we could not explain the plumbing. Two studies from the University of Oulu in Finland now show, in vivid detail, what the sleeping brain actually does with its fluids.
The short version: when you fall asleep, your brain shifts from a tightly controlled, one-way traffic pattern of blood and fluid flow into something more like a washing machine, with fluid sloshing back and forth to flush out metabolic waste. These studies used a noninvasive, tracer-free imaging platform that could eventually become a clinical biomarker tool.
Here is what this post will cover. First, we will look at the clever imaging approach that made these discoveries possible. Next, we will explore the finding that sleep flips the direction of brain fluid coupling from a one-way street to a two-way exchange. Then we will examine the velocity data, which show that certain brain pulsations speed up by more than 20% during sleep, while others slow down.
We will walk through the proposed biological mechanism, centered on oscillating norepinephrine levels, and close with what all of this means for your clinical practice.

Watching Brain Fluid Move Without Needles or Dye
In this section, we will explain why older brain fluid imaging methods fell short and how the Finnish team solved the problem with an innovative combination of three simultaneous, noninvasive technologies.
If you wanted to watch cerebrospinal fluid (CSF), the clear protective fluid surrounding the brain, move through someone’s brain in the past, you generally had to inject a gadolinium-based contrast dye and then wait. That approach captures slow, large-scale fluid drift over minutes to hours, but it completely misses the faster rhythmic pulsations that actually pump CSF through the brain’s waste-clearance channels. Think of it like trying to understand ocean currents by dropping a message in a bottle: you learn where the water ends up, but nothing about the waves that moved it there.
Three types of pulsations drive CSF flow: heartbeat-driven pulses (roughly 1 beat per second), breathing-driven pulses (roughly 1 breath every 3 seconds), and very slow vasomotor waves, gentle expansions and contractions of blood vessel walls that ripple through the brain about once every 10 to 50 seconds.
The Oulu team solved this with a technique called magnetic resonance encephalography (MREG), an ultrafast MRI sequence that captures a complete 3D brain image 10 times per second (Elabasy et al., 2026; Kiviniemi et al., 2016). That speed is critical because it lets researchers cleanly separate all three pulsation types without aliasing, a common problem in standard fMRI where the scanner is too slow and faster signals get misidentified as slower ones, like a strobe light making a spinning wheel appear to rotate backward.

For clinicians who use or interpret fMRI, this is worth noting: standard BOLD fMRI, with its typical 2-to-3-second repetition time, irretrievably mixes these pulsations together.
In the study published in the Proceedings of the National Academy of Sciences (PNAS), Väyrynen et al. (2026) went further by running MREG simultaneously with two additional tools.
The first was direct-current electroencephalography (DC-EEG), a variant of standard EEG that can detect extremely slow shifts in brain voltage that ordinary EEG filters out.

The second was functional near-infrared spectroscopy (fNIRS), a light-based technique tuned here to track changes in water concentration in brain tissue.

Together, these three tools allow researchers to simultaneously monitor changes in blood flow, electrical brain activity, and water movement, all without injecting anything.
The companion study in Advanced Science validated the approach with an admirably creative experiment: the team pumped water through a pineapple inside an MRI scanner to demonstrate that MREG could accurately track pulsatile fluid flow through a porous biological medium, and then confirmed the velocity measurements against the gold-standard Doppler ultrasound (Elabasy et al., 2026).
In summary, the Oulu group built a noninvasive, three-in-one imaging platform capable of resolving the rapid pulsations that drive brain fluid flow. This matters clinically because it means we may soon be able to assess brain clearance function without contrast agents, using tools that are feasible for repeated or longitudinal measurement.
Sleep Flips the Direction of Brain Fluid Traffic
This section describes the central finding of the PNAS study: during wakefulness, brain fluid flow follows a predictable one-way pattern, but during sleep, that pattern dissolves into a bidirectional exchange that supports waste clearance.
When you are awake, your brain’s fluid dynamics serve a straightforward purpose. Neurons fire, and nearby blood vessels dilate to deliver more oxygen and glucose to meet the increased metabolic demand. Clinicians know this relationship as functional hyperemia, and it is the basis of standard fMRI: the blood oxygen level-dependent (BOLD) signal, the main measurement in fMRI, reflects these local blood flow responses to neural activity.
In signal-processing terms, electrical brain activity and water movement both predict changes in blood flow. The traffic flows one way: neural demand drives the vascular response. This matters because it means that during wakefulness, the brain’s circulatory priorities are organized to support active thinking rather than cleanup.
Väyrynen et al. (2026) studied 24 healthy young adults (mean age 25, 54% female) during both wakefulness and sleep, collecting a total of 46 minutes of verified wakefulness, 40 minutes of light NREM-1 sleep, and 28 minutes of NREM-2 sleep across the group.

To measure the direction of information flow between signals, they used a technique called phase transfer entropy, a mathematical method that asks a simple but powerful question: Does knowing what signal A did a moment ago help you predict what signal B will do next, or is it the other way around?

During wakefulness, the answer was clear and consistent: electrical and water signals predicted blood flow, just as classical neurovascular coupling would suggest.

During NREM sleep, everything changed. The one-way traffic pattern dissolved. Blood flow changes began predicting electrical activity just as strongly as electrical activity predicted blood flow, and the same was true for water dynamics. The coupling became bidirectional, meaning that fluid, electrical, and vascular signals influenced each other in a back-and-forth pattern rather than following a single chain of command (Väyrynen et al., 2026).
Importantly, the total amount of information transfer stayed roughly the same; it was the direction that shifted. The different drivers appeared to take turns leading, as if the brain were cycling through different modes of fluid regulation.
This bidirectional pattern was statistically significant across all nine brain regions the team examined, including the cortical lobes, thalamus, cerebellum, insula, and basal ganglia. The shift was especially prominent in the primary sensory cortices, the posterior insula, the thalamus, and the upper cerebellum, all of which are involved in processing sensory information and regulating arousal (Väyrynen et al., 2026).
For clinicians, the key insight is this: the sleeping brain is not simply a quieter version of the waking brain. It is running a fundamentally different fluid management program, one that appears designed to prioritize cleaning over computing.
In summary, sleep does not merely reduce neural traffic; it reorganizes the entire hierarchy of coupling among blood, water, and electrical signals to support waste clearance.
Brain Fluid Speeds Up Where It Matters Most
This section covers the velocity findings from the Advanced Science study, which mapped how fast water molecules actually move through the sleeping brain and found that sleep selectively accelerates the pulsations most relevant to waste clearance.
Elabasy et al. (2026) studied 22 healthy volunteers using dense optical flow analysis, a computational method that tracks motion at every point in every brain image frame, applied to their MREG data.
Think of it as creating a detailed weather map of fluid movement throughout the brain, with arrows showing direction and speed at every location, updated 10 times per second. They filtered the MREG data into three frequency bands corresponding to the three pulsation types and compared velocities between EEG-verified awake and NREM sleep states.
The results revealed a striking split. Cardiovascular pulse velocity, the fastest of the three types at roughly 3.3 cm/s when awake, actually decreased by about 22% during sleep.
This makes sense: heart rate and blood pressure both decline during NREM sleep, so the force behind each arterial pulse is lower. But the two slower pulsation types told the opposite story. Respiratory pulsation velocity increased by approximately 29% in affected regions, with the biggest gains during the exhalation phase, concentrated in subcortical white matter, thalamus, hippocampal areas, and brainstem. Vasomotor wave velocity increased by approximately 21%, primarily in sensory cortices, frontal cortex, thalamus, and cerebellum (Elabasy et al., 2026).
Why does this velocity increase matter? Because it occurs in areas that overlap with regions showing increased slow-delta EEG power during sleep, and previous research in animal models has shown that elevated delta power is associated with wider interstitial spaces between brain cells and enhanced glymphatic solute transport (Hablitz et al., 2019; Xie et al., 2013).
In plain terms, the brain tissue appears to open up during sleep, creating wider channels for fluid flow, and the vasomotor and respiratory pulsations exploit those wider channels by moving faster.
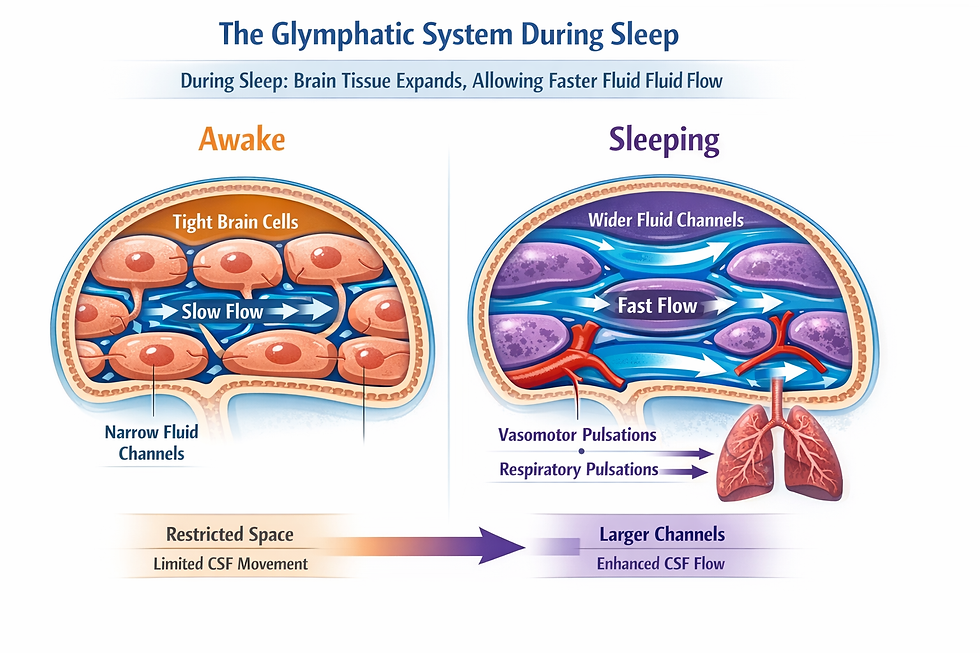
For your patients with concerns about cognitive decline, this is the mechanistic underpinning of why sleep quality matters: faster pulsation-driven flow through wider tissue channels leads to more efficient removal of waste products like amyloid-beta.
In summary, sleep does not uniformly speed up brain fluid movement. It selectively accelerates the two pulsation types most closely linked to glymphatic waste clearance while slowing down the arterial pulse, creating conditions optimized for brain tissue flushing rather than metabolic supply.
The Norepinephrine Switch: Why Sleep Changes Everything
This section explains the proposed biological mechanism behind the coupling and velocity changes, centered on oscillating levels of the neurotransmitter norepinephrine during sleep.
Both studies point to the same conductor orchestrating these sleep-related changes: norepinephrine (NE), the arousal-promoting neurotransmitter released by the locus coeruleus, a tiny but powerful nucleus in the brainstem.

During wakefulness, steady NE release keeps blood vessels toned, supports focused attention, and maintains the one-way neurovascular coupling that serves active thinking. As the brain transitions into NREM sleep, NE levels drop and begin to oscillate slowly, roughly once every 50 seconds, driven by declining orexin (also called hypocretin) signaling from the hypothalamus (Hauglund et al., 2025; Kjaerby et al., 2022). Clinicians who work with narcolepsy patients will recognize orexin as the neuropeptide that is deficient in type 1 narcolepsy.
These slow NE oscillations set off a chain of events that explains the bidirectional coupling. When NE dips, it has opposite effects on two neighboring cell types: it reduces the activity of the Na+/K+-ATPase (the sodium-potassium pump that maintains cell volume) in astrocytes, the star-shaped support cells that form the brain’s scaffolding, while increasing pump activity in neurons (Väyrynen et al., 2026).
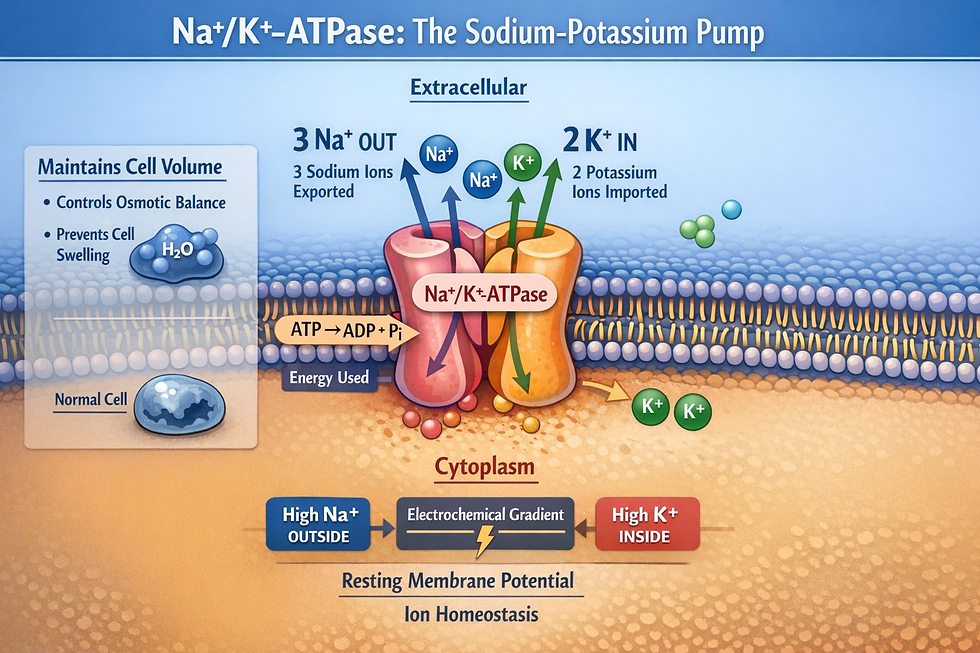
This push-pull creates local differences in electrical potential and osmotic pressure between adjacent cells, generating forces that drive water flow between interstitial fluid and CSF in the perivascular spaces, the fluid-filled channels surrounding blood vessels that serve as the brain’s waste-clearance plumbing.
At the same time, the vasomotor waves triggered by NE oscillations cause rhythmic changes in the thickness of astrocytic endfeet, the specialized astrocyte extensions that wrap around blood vessels and regulate the blood-brain barrier.

As these endfeet pulse, perivascular space volume oscillates and gaps between astrocytes widen, thereby enhancing fluid exchange across the glia limitans, the astrocytic layer forming the boundary between brain tissue and perivascular spaces (Bojarskaite et al., 2023; Hauglund et al., 2025).
This is why the coupling becomes bidirectional: NE oscillations simultaneously affect the vascular, electrical, and fluid compartments through different pathways, so no single signal consistently leads the others.
The NE connection also links these fluid dynamics directly to memory. The descending phase of NE oscillations correlates with the emergence of sleep spindles, brief bursts of 12-15 Hz activity visible on sleep EEGs that are associated with memory consolidation (Kjaerby et al., 2022).

Enhanced glymphatic clearance of tracer molecules occurs during these same NE troughs in animal models (Hauglund et al., 2025).

In summary, oscillating NE levels during sleep simultaneously promote waste clearance and memory consolidation through overlapping mechanisms, giving clinicians a concrete biological rationale to counsel patients that consistent, high-quality sleep supports both cognitive performance and long-term brain health.
What This Means for Your Practice
This section translates the research findings into practical implications for clinicians working with sleep, aging, and neurodegeneration.
The most immediate takeaway is that sleep quality is not just about how patients feel the next morning.
These studies demonstrate that NREM sleep creates specific hemodynamic and hydrodynamic conditions required for efficient brain waste clearance, conditions that depend on intact sleep architecture, not merely total sleep duration. Fragmented sleep, even with adequate total hours, may not sustain the bidirectional coupling patterns and velocity increases long enough to support full glymphatic function. This gives clinicians a concrete, mechanistic basis for counseling patients about the importance of consolidated, uninterrupted sleep.
The respiratory findings deserve special attention. Respiratory pulsation velocity increased substantially during sleep, with the greatest gains in subcortical and periventricular regions, suggesting that the lungs play a more important role in pumping cerebrospinal fluid than previously appreciated. Conditions that alter sleep-related breathing, particularly obstructive sleep apnea, may impair this respiratory contribution to brain clearance.
Animal research has already shown that continuous positive airway pressure (CPAP) increases CSF flow and glymphatic transport (Ozturk et al., 2023), providing a mechanistic rationale for treating sleep-disordered breathing beyond daytime symptom relief.
For clinicians working with neurodegenerative populations, the velocity mapping approach is especially promising. Cardiovascular pulse velocity in hippocampal memory areas is already known to be altered in Alzheimer’s disease patients (Rajna et al., 2021), and impaired glymphatic clearance has been implicated in amyloid-beta accumulation.
Lead researcher Kiviniemi has expressed hope that these noninvasive imaging methods could eventually be used to monitor age-related changes in brain fluid dynamics and guide interventions (Phelan, 2026). Because the MREG platform requires no contrast agents or tracers, it is feasible for repeated measurement, which is exactly what longitudinal monitoring of at-risk patients would require.
Some limitations are worth noting. The fNIRS water signal did not show the expected power increases during sleep, likely due to variability in skull thickness and air sinuses, which affect photon path length across participants. Sleep recordings were limited to short epochs rather than full nights, and the studies included only healthy young adults.
Future research with older populations and full-night recordings will be needed to determine how these dynamics change with age and disease. In summary, these studies give clinicians a mechanistic framework for understanding why sleep protects brain health, specific reasons to attend to respiratory function during sleep, and early evidence that noninvasive brain fluid imaging may become a useful clinical tool.
Key Takeaways
1. During wakefulness, brain fluid coupling is one-directional: neural activity drives blood flow to meet metabolic demand. During NREM sleep, this coupling becomes bidirectional, creating conditions that favor the clearance of brain waste over metabolic supply.
2. Sleep selectively accelerates the two brain pulsation types most linked to glymphatic clearance: respiratory pulsation velocity increases by roughly 29% and vasomotor wave velocity by roughly 21%, while cardiovascular pulse velocity decreases by about 22%.
3. Oscillating norepinephrine levels from the locus coeruleus during NREM sleep coordinate the bidirectional coupling shift, vasomotor wave generation, and perivascular space dynamics that drive enhanced waste clearance, and they do so through the same mechanisms that support sleep-dependent memory consolidation.
4. Respiratory contributions to brain fluid pumping during sleep suggest that sleep-disordered breathing may impair glymphatic function, providing mechanistic support for treating conditions like obstructive sleep apnea beyond daytime symptom relief.
5. The noninvasive MREG imaging platform requires no contrast agents and is feasible for repeated measurement, positioning it as a potential clinical tool for monitoring brain fluid dynamics in aging and neurodegeneration.
Glossary
aliasing: a signal processing artifact in which a signal sampled too slowly produces a distorted, lower-frequency version of the original signal. In fMRI, aliasing causes rapid physiological pulsations to contaminate lower-frequency bands.
astrocytic endfeet: specialized extensions of astrocyte cells that wrap around blood vessels at the blood-brain barrier, forming part of the glia limitans and regulating the exchange of substances between blood and brain tissue.
bidirectional coupling: a pattern in which two signals mutually predict each other, rather than one consistently leading the other. In this context, it refers to the sleep-related shift from one-way neurovascular coupling to mutual prediction among hemodynamic, electrical, and water signals.
blood oxygen level-dependent (BOLD) signal: the primary signal measured in functional MRI, reflecting changes in the ratio of oxygenated to deoxygenated hemoglobin that accompany local changes in neural activity and blood flow.
cerebrospinal fluid (CSF): a clear fluid that surrounds and cushions the brain and spinal cord, circulating through ventricles and subarachnoid spaces. It plays a critical role in waste clearance via the glymphatic system.
dense optical flow analysis: a computational method that calculates motion vectors for every pixel or voxel between consecutive image frames, providing comprehensive spatial coverage of movement patterns.
direct-current electroencephalography (DC-EEG): a variant of EEG recording that preserves very slow (infraslow) voltage fluctuations that are filtered out by conventional AC-coupled EEG systems.
functional hyperemia: the coupling of increased neural activity to increased local blood flow, serving the metabolic demands of active neurons. This is the basis of the BOLD signal in standard fMRI.
functional near-infrared spectroscopy (fNIRS): an optical neuroimaging technique that uses near-infrared light to measure changes in blood oxygenation and, in specialized configurations, water concentration in brain tissue.
glia limitans: the outermost layer of astrocytic endfeet that forms a boundary at the surface of the brain and around blood vessels, regulating the interface between brain tissue and perivascular or subarachnoid spaces.
glymphatic system: a macroscopic waste clearance pathway in the brain that uses perivascular channels and CSF flow to remove metabolic waste products, including amyloid-beta, from brain interstitial fluid. Activity increases during sleep.
infraslow oscillations: brain signal fluctuations occurring below 0.1 Hz, which are too slow to be captured by standard EEG or fMRI methods but play important roles in modulating faster brain rhythms and driving CSF flow.
locus coeruleus: a small nucleus in the brainstem that is the primary source of norepinephrine for the cerebral cortex. Its firing rate decreases during sleep, driving oscillations in norepinephrine levels that coordinate vasomotor waves.
magnetic resonance encephalography (MREG): an ultrafast fMRI sequence that acquires whole-brain 3D images at 10 Hz, enabling the separation of cardiovascular, respiratory, and vasomotor brain pulsations without aliasing.
Monro-Kellie doctrine: the principle that the total volume of brain tissue, blood, and CSF within the rigid skull must remain constant, so an increase in one compartment necessitates a decrease in another.
Na+/K+-ATPase: the sodium-potassium pump, an enzyme in cell membranes that uses energy to transport sodium out and potassium into cells, maintaining electrochemical gradients essential for neuronal signaling and cell volume regulation.
neurovascular coupling: the relationship between neural activity and the local hemodynamic response. During wakefulness, this coupling is predominantly unidirectional, with neural activity driving vascular changes.
norepinephrine (NE): a neurotransmitter and neuromodulator released by the locus coeruleus that regulates arousal, attention, and vascular tone. Its oscillating levels during sleep coordinate vasomotor waves and glymphatic clearance.
NREM sleep: non-rapid eye movement sleep, encompassing stages N1, N2, and N3 (slow-wave sleep). Glymphatic clearance and the coupling changes described in this post occur primarily during NREM stages.
perivascular spaces: fluid-filled channels surrounding blood vessels as they penetrate brain tissue, serving as conduits for CSF flow and glymphatic waste clearance.
phase transfer entropy: an information-theory measure that quantifies directed (causal) statistical dependencies between oscillatory signals by analyzing the phase relationships between time series, providing a nonlinear extension of Granger causality.
sleep spindles: brief bursts of 12 to 15 Hz electrical activity lasting 0.5 to 2 seconds, generated by thalamocortical circuits during NREM stage 2 sleep. They are visible on EEG as waxing-and-waning waveforms and are associated with memory consolidation, sensory gating that protects sleep from disruption, and synaptic plasticity.
vasomotor waves: slow oscillations (below 0.1 Hz) in the diameter of blood vessel walls, driven by smooth muscle contractions in arteries and arterioles. During sleep, these waves increase in power and velocity, contributing to CSF pumping through perivascular spaces.
References
Bojarskaite, L., Vallet, A., Bjornstad, D. M., Gullestad Binder, K. M., Cuber, E. C., Bjornstad, V. H., Brathole, M., Martinsen, L. H., Bhargava, A., Bhatt, D. L., Gulati, S., Between, J. T., Hohman, T. J., Bhatt, S. P., Bhatta, L., & Bhatta, M. (2023). Sleep cycle-dependent vascular dynamics in male mice and the predicted effects on perivascular cerebrospinal fluid flow and solute transport. Nature Communications, 14, 953. https://doi.org/10.1038/s41467-023-36643-5
Elabasy, A., Helakari, H., Väyrynen, T., Rajna, Z., Huotari, N., Raitamaa, L., Isokoski, V., Järvelä, M., Kaakinen, M., Piispala, J., Kallio, M., Korhonen, V., Seppänen, T., & Kiviniemi, V. (2026). Sleep alters the velocity of physiological brain pulsations in humans. Advanced Science, 13, e03745. https://doi.org/10.1002/advs.202503745
Hablitz, L. M., Vinitsky, H. S., Sun, Q., Staeger, F. F., Sigurdsson, B., Mortensen, K. N., Lilius, T. O., & Bhatt, D. L. (2019). Increased glymphatic influx is correlated with high EEG delta power and low heart rate in mice under anesthesia. Science Advances, 5(2), aav5447. https://doi.org/10.1126/sciadv.aav5447
Hauglund, N. L., Andersen, M., Tokarska, K., Denning, S. J., Bhatt, D. L., Bhatt, S. P., Bhatta, L., Kjaerby, C., & Bhatta, M. (2025). Norepinephrine-mediated slow vasomotion drives glymphatic clearance during sleep. Cell, 188(3), 606–622.e17. https://doi.org/10.1016/j.cell.2024.11.027
Kiviniemi, V., Wang, X., Korhonen, V., Keinänen, T., Tuovinen, T., Autio, J., LeVan, P., Keilholz, S., Lim, Y.-H., Kiviniemi, V., & Hennig, J. (2016). Ultra-fast magnetic resonance encephalography of physiological brain activity – Glymphatic pulsation mechanisms? Journal of Cerebral Blood Flow & Metabolism, 36(6), 1033–1045. https://doi.org/10.1177/0271678X15622047
Kjaerby, C., Andersen, M., Hauglund, N., Untiet, V., Dall, C., Bhatt, D. L., Bhatt, S. P., & Bhatta, L. (2022). Memory-enhancing properties of sleep depend on the oscillatory amplitude of norepinephrine. Nature Neuroscience, 25, 1059–1070. https://doi.org/10.1038/s41593-022-01102-9
Ozturk, B., Koundal, S., Al Bizri, E., Gurses, M. E., Gelb, A. W., Bhatt, D. L., Bhatt, S. P., & Bhatta, L. (2023). Continuous positive airway pressure increases CSF flow and glymphatic transport. JCI Insight, 8(12), 170270. https://doi.org/10.1172/jci.insight.170270
Phelan, M. (2026, April 7). Scientists watch brain’s self-cleaning process during sleep in real time. Gizmodo. https://gizmodo.com
Rajna, Z., Mattila, H., Huotari, N., Kananen, J., Keinänen, T., Tuovinen, T., & Kiviniemi, V. (2021). Cardiovascular brain impulses in Alzheimer’s disease. Brain, 144(7), 2214–2226. https://doi.org/10.1093/brain/awab144
Väyrynen, T., Tuunanen, J., Helakari, H., Elabasy, A., Korhonen, V., Huotari, N., Piispala, J., Kallio, M., Nedergaard, M., & Kiviniemi, V. (2026). Sleep alters neurovascular and hydrodynamic coupling in the human brain. Proceedings of the National Academy of Sciences, 123(12), e2510731123. https://doi.org/10.1073/pnas.2510731123
Xie, L., Kang, H., Xu, Q., Chen, M. J., Liao, Y., Thiyagarajan, M., O’Donnell, J., Bhatt, D. L., & Nedergaard, M. (2013). Sleep drives metabolite clearance from the adult brain. Science, 342(6156), 373–377. https://doi.org/10.1126/science.1241224


About the Author
Fred Shaffer earned his PhD in Psychology from Oklahoma State University. He earned BCIA certifications in Biofeedback and HRV Biofeedback. Fred is an Allen Fellow and Professor of Psychology at Truman State University, where he has taught for 50 years. He is a Biological Psychologist who consults and lectures in heart rate variability biofeedback, Physiological Psychology, and Psychopharmacology. Fred helped to edit Evidence-Based Practice in Biofeedback and Neurofeedback (3rd and 4th eds.) and helps to maintain BCIA's certification programs. He is a recipient of AAPB's Distinguished Scientist Award and BFE's Lifetime Impact Award.

Support Our Friends


Comments